
Động vật không xương sống, trong đó có tôm chỉ phát triển hệ miễn dịch tự nhiên (innate immunity). Hệ miễn dịch tự nhiên chia ra làm hai hệ thống bảo vệ chính: miễn dịch tế bào (cellular barriers) và miễn dịch dịch thể (Humoral barriers). Miễn dịch tế bào bao gồm hệ thống thực bào (phagocytocis), tạo thể bao (encapsulation), thể hạch (nodulation) bao vây vật thể lạ xâm nhập và sau đó phá hủy thông qua hệ thống hoạt hóa Pro-phenoloxidase (proPO activating system). Ngoài ra, trong đáp ứng miễn dịch kháng virus còn có cơ chế apoptosis. Miễn dịch dịch thể bao gồm các hệ thống: bổ thể (complement system), đông máu (hemolymph coagulation system), yếu tố dính kết tế bào (agglutinin – lectin system) và quá trình tổng hợp các loại peptide kháng khuẩn, kháng nấm và kháng virus. Ngoài ra, vỏ kitin ở tôm là rào cản vật lý có vai trò trong các quá trình sinh lý khác nhau liên quan đến các phản ứng miễn dịch.
1. Đáp ứng miễn dịch tế bào
1.2. Thành phần tế bào
Động vật giáp xác có hệ thống tuần hoàn mở với máu màu xanh lá cây, tuần hoàn qua mạch máu và đưa đến các mô. Các tế bào máu (hemocytes) và thành phần dịch thể được vận chuyển trong máu giúp chúng tiếp xúc dễ dàng với các phân tử từ bên ngoài. Sự tạo máu hình thành nên các tế bào trưởng thành của hệ thống miễn dịch tự nhiên, số lượng tế bào máu thể hiện vai trò bảo vệ vật chủ và sự cân bằng. Sự hình thành tế bào máu được điều khiển bởi các tín hiệu bên trong và ngoài tế bào, kết quả dẫn đến sự hoạt hóa của hàng loạt tín hiệu đặc trưng. Mô tạo máu (hematopoietic tissue – HPT) ở động vật giáp xác là một mạng lưới của tiểu thùy nằm ở hai bên lưng và mặt trên của dạ dày, gần động mạch dâu và ở gốc hàm. Các tế bào máu được tạo ra trong thành của các ống này và giải phóng vào trong các khoang mạch. HPT của tôm sú và tôm penaeid khác nằm trong các vùng khác nhau của dạ dày, hàm và tuyến râu.
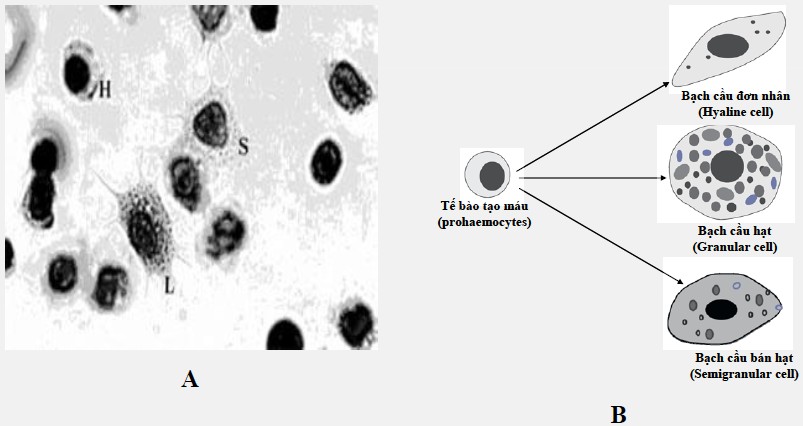
Hình 1. Tế bào máu tôm sú. A. Tế bào máu tôm sú chụp dưới kính hiển vi: H (Hyaline cell – Bạch cầu đơn nhân; S (Semigranular cell – bạch cầu bán hạt); L (Large granular cell – Bạch cầu hạt), B. Mô phỏng các loại tế bào máu ở giáp xác
Các tế bào máu ở họ tôm Penaeid nói chung và ở tôm sú nói riêng có đặc tính sinh học và chức năng tương tự với các đại thực bào, bạch cầu hạt và tế bào giết tự nhiên của động vật có xương sống. Những tế bào này thực hiện các quá trình thực bào, hình thành thể hạch, thể bao, melanin, hoạt hóa proPO và apoptosis. Chúng cũng tạo ra các yếu tố dính kết tế bào (agglutinin-lectin system), hệ thống đông máu (hemolymph coagulation system), hệ thống bổ thể (complement system), và peptide kháng khuẩn (AMP). Các tế bào máu giải phóng các enzyme ức chế cần thiết để điều khiển phân giải nhiều protein, ngăn chặn sự biểu hiện quá nhiều protein và các mô bị phá hủy.
Có ba loại tế bào máu (hemocytes) ở tôm: bạch cầu đơn nhân, bạch cầu hạt và bạch cầu bán hạt (Hình 1). Các bạch cầu đơn nhân chiếm 5 – 15% tế bào máu lưu thông, là những tế bào nhỏ không khúc xạ, có nhân nhỏ, trong tế bào chất có ít hoặc không có hạt tế bào chất. Bạch cầu đơn nhân không có hoạt động thực bào và dễ dàng bám vào các bề mặt kính, giống như các đại thực bào ở cá và động vật có vú. Vai trò chính của các tế bào này có liên quan đến quá trình đông máu và thực bào. Bạch cầu hạt chiếm 10-20% tế bào máu lưu thông, nhân nhỏ nhất trong số các tế bào máu. Loại tế bào này có thể bị kích thích bởi β-1,3-glucans, peptidoglycans (PG) và lipopolysaccharides (LPS) để gây nên sự xuất bào (exocytosis) và giải phóng enzyme. Ngoài ra, trong tế bào chứa lượng lớn hạt tế bào chất (hạt có chiều rộng 0,8uM), các hạt sẽ giải phóng ra các protein bám β-glucan; lipopolysaccharide; peptidoglican, các yếu tố đông máu (transglutaminase), các yếu tố liên quan đến enzyme prophenoloxidase (proPO; ppA; peroxinectin), các chất ức chế proteinase (alpha-macroglobulin; karazl; serpin; pacifastin) và các chất kháng khuẩn (penaeidin; lectin). Chức năng của chúng là hình thành thể bao, khởi động hệ thống proPO và thực bào. Các bạch cầu bán hạt chiếm 75% tế bào máu lưu thông, có một số lượng lớn các hạt nhỏ (hạt có chiều rộng 0,4 micromet) giống như bạch cầu hạt ở động vật có xương sống. Các tế bào này chứa các thụ thể cho β-1,3-glucans và hoạt động chức năng chủ yếu của chúng liên quan đến thực bào, tạo thể bao để bao bọc các vật thể lạ và đông máu.
1.3. Các quá trình đáp ứng miễn dịch tế bào
Thực bào là cơ chế miễn dịch tế bào chính ở tôm liên quan đến sự thu nhận các vật thể lạ từ bên ngoài. Cơ chế này được thực hiện bởi bạch cầu bán hạt và bạch cầu hạt. Quá trình thực bào bao gồm các bước: hướng hóa, bám chặt, tiêu hóa, phá hủy tác nhân gây bệnh và xuất bào. Nhìn chung, quá trình thực bào ở động vật không xương sống nói chung và tôm nói riêng tương tự như ở động vật có xương sống. Tuy nhiên, sự tiếp xúc miễn dịch luôn được thực hiện có tính ngẫu nhiên, rất hiếm chủ động theo kiểu hóa hướng động ở động vật có xương sống. Ngoài ra, bạch cầu bán hạt có khả năng nhận biết các tác nhân xâm nhập và hoạt động như là một sự opsonins hóa kết hợp với hệ thống hoạt hóa proPO. Các thể hạch phân giải tế bào máu phát hiện trong gan tụy và mang, được hình thành bởi nhiều tế bào máu cùng hoạt động hỗ trợ để bẫy các vi sinh vật hoặc các kháng nguyên lớn mà không thể loại bỏ bởi cơ chế thực bào. Sau đó, các thể hạch này trải qua sự hoạt hóa của hệ thống proPO, sự melanin hóa và phá hủy vi khuẩn.
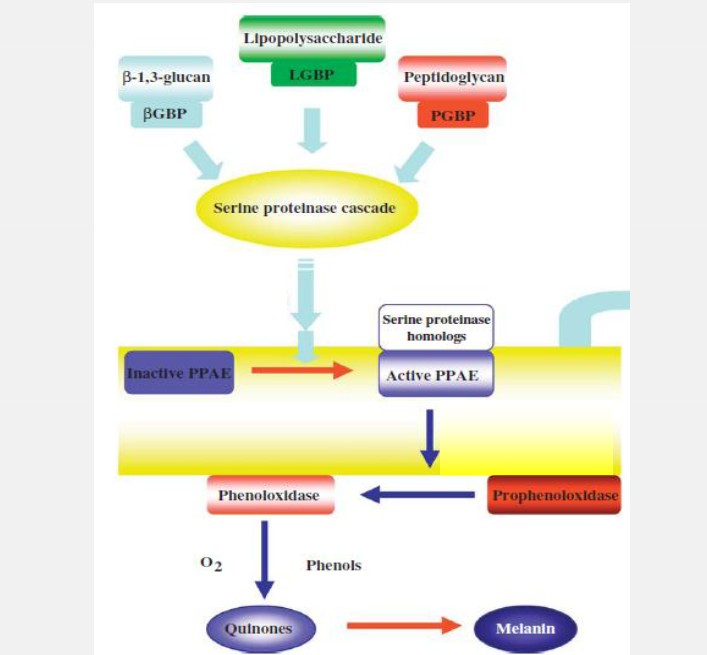
Hình 2. Hệ thống hoạt hóa proPO và tổng hợp melanin
Các yếu tố chống oxy hóa bảo vệ tôm khỏi những tác động gây độc tế bào gây ra bởi trao đổi chất tế bào và stress oxy hóa tạo ra bởi sự mất cân bằng của các chất phản ứng ôxy trung gian (reactive oxygen intermediates – ROIs) và chất phản ứng nitơ trung gian (reactive nitrogen intermediates – RNIs). Để ngăn chặn sự phá hủy này, chiến lược bảo vệ chất chống oxy hóa đã được phát triển bao gồm các enzyme (catalase, glutathione peroxidase (GPX), SOD) và các thành phần khác (ascorbate, β-carotene, flavonoid, α-tocopherol và vitamin E) có thể trung hòa ROIs hoặc sửa chữa sự phá hủy ở mức độ phân tử cho tế bào. Bằng thực nghiệm Down và đtg (2001) đã đưa ra kế t luậ n các enzyme chống oxy hóa đóng vai trò như các chất điều khiển đáp ứng miễn dịch. Hoạt động của enzyme superoxide dismutase (SODs) là một trong những cơ chế bảo vệ chính chống lại stress oxy hóa gây ra bởi ô nhiễm, lây nhiễm bệnh, tình trạng thiếu oxy (hypoxia), quá nhiều oxy (hyperoxia), nhiệt độ và các chất kích thích miễn dịch. Một phân tử SOD ngoại bào (EC-SOD) đã được công bố ở tôm hùm (Homarinus capensis), nó kết hợp với sự thực bào, sự hình thành thể bao, sự opsonin hóa và tạo ra các hợp chất diệt khuẩn. Tuy nhiên, việc tạo ra các hợp chất oxy hóa có tác dụng kháng khuẩn đã được nghiên cứu trong các tế bào máu ở động vật không xương sống. Sự đáp ứng tế bào của các chất oxy hóa là rất nhanh và được tạo ra trong suốt quá trình thực bào vi khuẩn. Những hợp chất này bao gồm anion superoxide (O2-), gốc tự do hydroxyl (OH-), H2O2, ROIs và RNIs.
Bạch cầu hạt tổng hợp, lưu trữ và bài tiết hệ thống prophenoloxydase (proPO) được hoạt hóa bởi β-glucans nấm, PG và LPS. Các phân tử này cảm ứng bạch cầu hạt tiết proPO không hoạt động và chuyển đổi chúng thành proPO enzyme. Enzyme này oxy hóa phenol thành quinon, có thể giúp tiêu diệt mầm bệnh và được sử dụng để tạo ra melanin. Ngoài ra, các protein tương tác với các thành phần trong quá trình proPO có liên quan với sự nhận biết và sự trao đổi của các tế bào máu (Hình 2).
Melanin hóa là một quá trình hóa sinh phức tạp, ít được biết đến, có liên quan tới các protease khác nhau (trypsine như serine, serine protease) có sự tham gia của hệ thống phenoloxidase. Sự melanin hóa đóng một vai trò quan trọng trong các cơ chế bảo vệ động vật không xương sống, trong đó một bao dày của hắc tố melanin được tạo ra xung quanh tác nhân từ bên ngoài. Melanin, một sản phẩm của hệ thống proPO, là một sắc tố màu nâu tối với đặc tính kháng khuẩn. Đặc tính diệt khuẩn của melanin và các tác nhân khác như O2-, các gốc hydroxyl được tạo ra trong quá trình hình thành quinon.
Lectin là một họ glycoprotein hoặc protein có một hoặc nhiều vùng chức năng, nhưng có chung 1 vùng bám carbonhydrat. Lectin được sắp xếp thành nhiều loại khác nhau. Chức năng của lectin có thể tham gia vào hàng rào đầu tiên chống lại nguồn bệnh, điều khiển miễn dịch và ngăn chặn sự tự miễn. Kuhlman và đtg (1989) cho rằng lectin có vai trò như là một protein bổ thể làm tăng quá trình thực bào. Ở tôm sú, lectin đã được tạo dòng và xác định trình tự.
Apoptosis là một quá trình tế bà o chế t theo chương trình xả y ra ở các cơ thể sinh vật đa bào và được bảo thủ qua tiến hóa. Nó đóng vai trò quan trọng trong duy trì hoạt động bình thường của nhiều mô và cơ quan ở cơ thể sinh vật đa bào bằng cách loại bỏ các tế bào đã bị phá hủy, đặt không đúng chỗ hoặc trở nên không cần thiết. Apoptosis đóng vai trò quan trọng trong các quá trình kháng virus ở nhiều sinh vật bằng cách loại bỏ những tế bào nhiễm virus. Do đó, apoptosis như là một cơ chế đáp ứng miễn dịch tự nhiên để hạn chế sự sinh sản virus và có thể làm giảm hoặc loại bỏ sự lây lan của virus trong cơ thể vật chủ. Tuy nhiên, nhiều virus đã có các gen mã hóa protein có khả năng ngăn chặn hiệu quả hoặc trì hoãn quá trình apoptosis để sinh sản với số lượng lớn. Một số virus có gen mã hóa protein có thể ức chế vào các caspase – protein trung tâm trong hoạt động của apoptosis. Mặt khác, số lượng virus tăng dẫn đến apoptosis hoạt động mạnh. Quá trình apoptosis có thể hoạt động như là một bước cuối cùng và quan trọng trong sự lây truyền virus đến tế bào lân cận. Số lượng các phân tử DNA của virus tăng trong tế bào máu dẫn đến hiện tượng apoptosis ở tôm nhiễm WSSV hoặc YHV. Các caspase là protein trung tâm trong quá trình apoptosis. Ở tôm thẻ Nhật Bản (Marsupenaeus japonicus) nhiễm WSSV gen caspase bị „im lặng‟, apoptosis bị ức chế đáng kể dẫn đến sự gia tăng các bản sao của WSSV, điều này chỉ ra rằng có thể apoptosis đóng vai trò trong cơ chế kháng virus ở tôm nuôi. Tỷ lệ các tế bào máu thực hiện apoptosis trong tôm nhiễm WSSV là rất thấp, nhưng cao hơn đáng kể hơn so với tôm càng tiêm WSSV vào ngày 3 hoặc 5 sau nhiễm. Apoptosis cũng được cho là có vai trò loại bỏ TSV ở tôm trong tự nhiên. Ở tôm sú, gen mã hóa caspase cũng đã được tạo dòng và xác định trình tự.
2. Đáp ứng miễn dịch dịch thể
Ngoài chức năng tham gia hoạt động trong sự opsonin hóa, thực bào, ngưng kết và đóng gói tác nhân gây bệnh, các lectin ở động vật không xương sống còn được coi là phân tử nhận biết nguyên thủy có khả năng phát hiện carbohydrate, thúc đẩy hệ thống hoạt hóa proPO. Các protein nhận biết mô hình (PRP) là lectins đã phát hiện các phân tử như LPS, PG, lipoteichoic acid của vi khuẩn, β-1,3-glucans ở nấm và RNA virus giúp hoạt hóa các cơ chế bảo vệ vật chủ. Các chức năng sinh học của PRPs là khởi đầu của sự hình thành một loạt các protein hoặc dấu hiệu của cơ chế bảo vệ và loại bỏ những tác nhân xâm nhiễm hệ thống máu. Khi PRPs phát hiện kháng nguyên, các tế bào máu di chuyển đến vị trí của chúng bằng hướng hóa, tạo ra một phản ứng viêm. Kết quả là một cơ chế bảo vệ nhanh và hiệu quả chống lại tác nhân gây bệnh. Thụ thể Toll receptor (TLRs) cũng là một họ PRRs cổ xưa có ở các động vật khác nhau có thể phát hiện tất cả các loại tác nhân gây bệnh. TLRs được hoạt hóa bởi sự lây nhiễm vi khuẩn và virus và đã được công bố ở tôm thẻ Trung Quốc và tôm thẻ chân trắng. Ở tôm sú, một số protein được cho rằng có thể đóng vài trò là thụ thể nhận biết mô hình: Rab7, Cyclic AMPregulated protein, kazal-type proteinase.
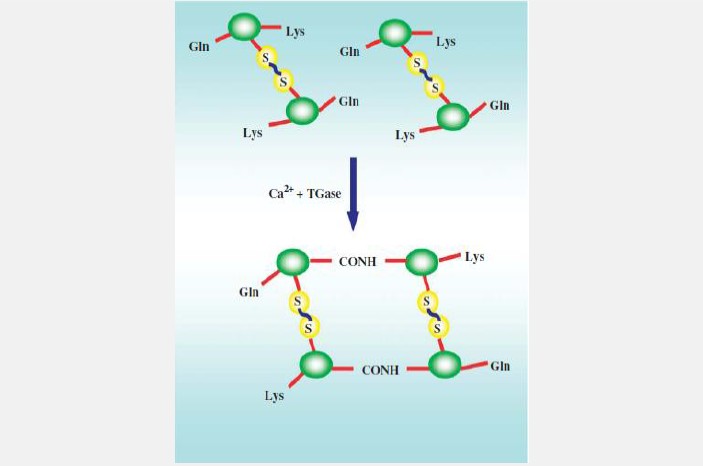
Hình 3. Cơ chế đông máu ở tôm
Quá trình đông máu được sử dụng để ngăn chặn sự mất máu do tổn thương ở vỏ kitin và làm bất động các tác nhân gây bệnh xâm nhập. Ở động vật giáp xác, quá trình đông máu được điều khiển bởi các protein đông máu (Hình 3). Các protein đông máu trong huyết tương hình thành nên dạng polymer cộng hóa trị nhờ transglutaminase phụ thuộc Ca2+ được tiết bởi các tế bào máu. Các protein đông máu của tế bào có thể được hoạt hóa bởi LPS hoặc β-1,3-glucan và có liên quan đến hệ thống hoạt hóa proPO. Đáp ứng miễn dịch dịch thể còn có sự tham gia của các phân tử như cytokine có khả năng hoạt hóa các đáp ứng kháng khuẩn; peroxynectin thực hiện chức năng phân tán tế bào máu, thực bào, hình thành thể bao, thể hạch và ngưng kết mà kết quả là sự hoạt hóa peroxide và tiêu hủy tác nhân xâm nhiễm; Protein sốc nhiệt (heat shock protein -HSP) hoặc chaperonins ở động vật không xương sống có khả năng bảo vệ và phục hồi cấu trúc của protein. Năm 2004, mối liên hệ giữa các protein sốc nhiệt, stress và các đáp ứng miễn dịch của tôm đã được thực hiện bởi Lo và đtg, nhóm nghiên cứu đã tách dòng và mô tả cDNA của gen HSP70 ở tôm sú. Phân tử HSP70 gồm 652 amino acid với khối lượng phân tử khoảng 71,4 kDa. Sự biểu hiện mRNA của HSP70 ở tế bào máu tôm tăng 2 đến 3 lần sau một giờ sốc nhiệt và trở lại trạng thái bình thường sau 30 phút.
Một yếu tố quan trọng chống tác nhân gây bệnh ở động vật không xương sống là các peptide kháng khuẩn (Antimicrobial peptides – AMPs). Đây là những protein có trọng lượng phân tử thấp, cần thiết ở các cơ thể sinh vật thiếu khả năng miễn dịch thích ứng. AMPs có phổ hoạt tính rộng, sự đặc hiệu thấp và gây độc yếu đối với tế bào động vật. Những peptide này tác động lên màng tế bào của vi khuẩn, nấm, ký sinh trùng, màng bao bọc virus và thậm chí cả tế bào ung thư, gây nên sự bất ổn định của các ion và năng lượng dẫn đến sự hình thành các lỗ trên màng tế bào. Lysosome là một thành phần tham gia vào quá trình thực bào, trong lysosome có chứa nhiều enzyme gọi chung là lysozyme. Lysozyme có vai trò làm giảm các polysaccharide ở màng nhày của thành tế bào vi khuẩn Gram (-), thay đổi cấu tạo phân tử của bề mặt tế bào, cho phép các tế bào thực bào dễ dàng nhận biết chúng. Hầu hết các lysozyme tham gia vào sự phá hủy các vi sinh vật bên trong và bên ngoài tế bào máu, một số còn đóng vai trò của sterases và chitinases.
Theo AquanetViet












